����̒�����������
(4)���茧�ɂ�����h���A�����`�̐����n
(5)�^�P�m�R�J���j�i�̖k���Y�����ɂ����镪�z
(7)�L�C���c�`�g�����` �������ێs�����ɂĔ���
(8)�V�I�A�����{�ƃV���E�~�A�����{���茧�����ێs��\�㓇�Ŋm�F����
(10)���茧�����ێs�Ɩk���Y�S���m�����̃��}�A�J�K�G���ƃj�z���A�J�K�G���̕��z�ɂ���
(11)�ܓ�����j�z���~�c�o�`��������
(12)���c���n�Ƃ��钷�茧�����ێs���m�����̃~���}�A�J�l�̌Q�̊ώ@�L�^
(13)���茧�k���Y�����y�ѕ��˓��̉͌���ɂ�����g���ނ̕��z�ɂ���
(14)���}�A�J�K�G���̐V�Y�n
(15)�T�̎q���̃q���n�l�� (384KB)
(16)2014�N�~���}�A�J�l�̕ۑS��
(17)�I�L�q���V�C�m�~�K�C�̎Y���L�^
(18)�F�v���̃V�}���N�Z�C
(19)�k���Y�����̐A���ώ@�L�^(9)
(20)�k���Y�����̃^�C�����A�I�C�̕��z
(21)���茧�ɂ�����c�V�}�P�}�C�}�C�ƃi�J�_�`�M�Z���̐V�Y�n
(22)���茧���L�^�̃��x�J�����`
(22)�k���Y�����̐A���ώ@�L�^�i10�j (612KB)
(23)���X��̃A�u���{�e�ƎY����L (80KB)
��J���
�@���\�㓇�ɖʂ���������̐����ɏ����Ƃ�������������]������܂��B���s������100m�A��10m���܂�ŁA���삩�狷������ʂ��ĎԂł��s�����Ƃ��ł��܂��B���̓���]�͐̂ő咪�̊������ɂ͘p���͊��オ��A�Ί݂̌��m���܂œn�ꂻ���Ȃ��炢�ł��B�u�����ێs���b�h�f�[�^�u�b�N2002�v�̒����̍ۂɁA�Óc�搶���炱�̏ꏊ���Љ��Ă͂��߂Č��ɗ��܂������A����]�̗��݂ɂ͍L�t�������Â��قǖ��Ă���A���Ƃ����\�����҂�����̕��͋C�ł����B�����ł܂��ڂɕt�����̂̓J�j�����ŁA���オ�������D�̋߂��ő����̃n�N�Z���V�I�}�l�L��`�S�K�j���������Ă��܂����B���̌�A�����̂��߂ɉ��x���K��܂������A���邽�тɐV��������������܂��B
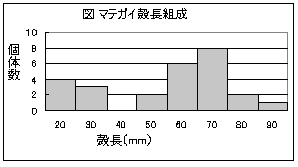
�@��N4��14���Ɋ����ׂĂ݂܂����B�����`�̕��`�g��݂��āA2m2�̖ʐς�[��30cm�قǃX�R�b�v�Ō@���āA�D�Ƃ�������ɏo�Ă����������̏W���Č���ڂ������ׂ܂����̂ŁA���̌��ʂ��Љ�܂��B
�@���������ʐ�2m2�̊����ɂ́A�L�ł̓}�e�K�C26�́A�V�I���K�C37�́A�J�K�~�K�C3�́A�I�L�V�W�~3�́A�A�T���̒���2�́A�����čb�k�ނł̓X�i���O���̒��Ԃ�85�̐������Ă��܂����B���̂����V�I���K�C�͍����ێs���b�h�f�[�^�u�b�N�Ő�Ŋ뜜�TB�ނɁA�܂��}�e�K�C�͓�������Ŋ뜜�U�ނɎw�肳��Ă����ł����B�A�T���̒��Ԃ̓q���A�T���̂悤�ł�����܂ł͂킩��܂���ł����B�X�i���O���̒��Ԃ���܂ł킩��܂���ł����B
�p���ɂ͌͂�t�������Ă��āA�����ڂł͕x�h�{�����Ă���悤�ł������A�C���̗��ʂ��悢���߂��A���܂�Ҍ�������Ă��܂���ł����B��������2m2�̊����ɐ������Ă��������̏d�ʂ͍��v683g�ł����B�����1 m2������340g�̓L��b�k�ނ��������Ă��āA�����̐������C�����̃v�����N�g������߂��A���a���Ă����L�@����H�ׂĂ���Ƃ������ƂɂȂ�܂��B
�͓̂��p�̕l�ɂ͂�������̃}�e�K�C�������̂ɁA�ߍ��߂����茩�Ȃ��Ȃ�܂������A�����ł͌��݂ł����B
�����ō̏W�����}�e�K�C�̃T�C�Y�ʂ̌̐���}�Ɏ����܂��B
�@���̐}������ƁA�k�̒���(�k��)��60�`80mm�̔�r�I�傫���L�������̂��킩��܂����A40mm�����̏������L�����邱�Ƃ���A�����ł͔ɐB�����}�e�K�C�����ꂵ�Ă��čĐ��Y���Ă��邱�Ƃ��킩��܂��B������������ł��B
 �@
�@
�����ɂ͂��̂ق��ɂ��J�j�ނƂ��ăn�N�Z���V�I�}�l�L�A�`�S�K�j�A�A�J�e�K�j�A���}�g�I�T�K�j�A�}���R�u�V�A�t�^�o�J�N�K�j���������Ă��܂��B����ɊL�ނł̓c���^�K�C�A�e���O�j�V�A�i�K�j�V�A�T���{�E�A�A�T���A�o�C�A�I�I�w�r�K�C�A�n�{�E�L�A����ɞ��瓮���̃��c�A�i�J�V�p���A�g�Q���~�W�Ȃǂ������܂��B�܂��A�Ă̏I��荠�ɂ͍b���̕������Z���`�Ɉ�����^�C�����K�U�~(���^���K�j)�̒t�K�j���A�������ۂ��j���ł��܂��B
2�N�O�܂ŁA�����̓�����ɂ��銱���ɂ̓R���c�L�K�j�������������Ă��܂������A��ݍH���ɂ�花�����Ȃ��Ȃ������߂ɃR���c�L�K�j�����ł��܂����B�܂������͐ς��Ă����̂ŕ������邩������܂��A�c�O�Ȃ��Ƃł��B����]�̉��͂����Ԑ̂ɂ��łɖ��ߗ��ĂĂ���A�Б��͓��H�ɂȂ��Ă��܂��B����ł������ɂ́A�l�̎�̓����Ă��Ȃ����R�̂܂܂̊C�݂�����\�㓇�̖ʉe������A��ɂ����������ۂ̎��R���̂ЂƂ��Ǝv���܂��B
(��J���)
�@��J���@�@�@�@�@�@
�@���Y��͉B���x�t�߂������Ƃ��鍲���ێs���ł͂����Ƃ�������ŁA��\�㓇�ɖʂ���劃���ɒ����ł��܂��B�͌��ɂ͍L��Ȋ������`������Ă��āA�N���}�G�r��K�U�~�i���^���K�j�j���͂��߂��܂��܂ȊL�ނ��������Ă��܂��B���̒��ɂ̓n�}�O�����܂������c���Ă��܂��B�n�}�O���͒W���̉e��������p�̔�r�I�ׂ������̊����ɐ������Ă��܂����A���̂悤�Ȋ����͖��ߗ��Ă̑ΏۂɂȂ�₷���A�܂��A�e��̔r���̉e���������Ƃ�������Ƃ���ł�����܂��B�܂��A�A�T���Ȃǂɔ�ׂāA�����⍻�̗��x�Ȃǂ̍D�ޔ͈͂��������߁A�e�n�Ō������Ă���Ƃ����b���܂��B���������āA���Y��̉͌��̃n�}�O���͑S���I�ɂ��M�d�Ȃ��̂��Ƃ��������ł��B
�@���̊����ł́A�咪�̊��������ɂ͉Ƒ��A��Ȃǂő����̐l�o������A�n�}�O�����n�߃J�K�~�K�C��V�I�t�L�A�o�J�K�C�Ȃǂ��̂��Ă��܂��B���ɂ�2cm���x�̏������n�}�O���������A��l�����āA���������^���܂��B�@4�����{�ɖ�1���Ԃ����č̏W���s���Ē��ׂ��n�}�O��86�̂̊k�̒����̑g�������}�Ɏ����܂��B�������k�̒����i�k���j�ŁA�c�����A���̃T�C�Y�̌̐��ł��B���������ƁA2cm���炢�̊L�����4cm���z���L�͂قƂ�ǂ��܂���B���̓��͊������Ɍv�������Ƃ���178�l�̐l�o���ώ@����܂����B���̂悤�Ȑl�o���咪�̖��ɉ����������ƍl����ƁA�̌Q�̃T�C�Y���������Ȃ��Ă���̂́A�߂肷���ɂ����̂��ƍl����ꂻ���ł��B
�@�n�}�O�����͂��߂Ƃ���L�́A�A���v�����N�g���̌]���Ȃǂ��h���Ƃ��ĐH�ׂĂ���A���̂�ߗʂ͖c��ȗʂł���Ƃ�������������܂��B�L�͐l�Ԃ̊����ɂ�萶�܂ꂽ�ߏ�ȉh�{���������ڂ��ʂ����Ă���Ƃ����Ă���A��ɂ��������̂ł��B
�@�����m�̕��������Ǝv���܂����A���N�O�܂ł͍��X��̉͌��̊����ł���ʂ̃n�}�O�����̂�Ă��āA�����̐l�łɂ�����Ă��܂������A�̂�s�����Ă��܂����̂��A�ߍ��ł͂قƂ�ǂ��Ȃ��Ȃ��������ł��i�ŋ߂킸���ɉ��Ă���Ƃ����b������܂��j�B������̐������͓��ɕς���Ă��Ȃ��悤�Ȃ̂ŁA���l�Ƃ����l�����܂���B�Z���Ԃɍ̂�s�����Ă��܂��̂ł͂Ȃ��A���������ĊL�@�肪�y���߂�悤�l���˂Ȃ�܂���B���̂��߂ɂ́A�n�}�O��������炵���l���ɂ���ƂƂ��ɁA�Y�����������Ȃ̂��A���V�c�����ǂ̂悤�ɒ��ꂵ�Ă���̂��A���ꂵ���t�L�̐��c���͂ǂ̂��炢�Ȃ̂��ȂǁA�����̊C��ɂ�����n�}�O���Ȃǂ̐����Ɋւ����b�����Ə����W���K�v���Ɗ����Ă��܂��B
���茧�����w��@��55�C2002
�����P���@
���茧�ɂ�����h���A�����`�̐����n�@
���茧�����w��@��55�C2002
�����P��
�h���A�����`�i�h���A�����`�ȁj�@Onchidium hongkongense Britton

�̒��R�p�A�̍�1.2�p�B���ȉ~�`�Ŗc����_�炩���B�O���w�ʂ͍��F�ŕs�K���Ȉ�Ȕ��������A�ア�C�{��̓ˋN������A��_�����݂���B���ĂȐG�o���Ȃ��B���͍����F�ŁA���ʂ̂قƂ�ǂ��߂�B
�����m���͘a�̎R���ȓ�C���{�C���͒��茧�Δn�ȓ�C�����C�z���R�����o�ăV���K�|�[���܂ŕ��z�B�傫�ȓ��p�̉��̕��R�ȍ��D�����ɂ��ށB�������͊����\�ʂ����C�����\�ʂ̑��ނ����ׂ�B�������͐[�����\cm�܂ō��D�̒��ɐ��荞�ނ��Ƃ�����B���R��w�_�w���̕��c�G�������ɂ��A��B�Ȗk�ł͂��Ēm��ꂽ�Y�n�̑����ł��łɐ�ł��C���ݎY�n�͌��ݐ��ӏ������m���Ă��Ȃ��Ƃ̂��Ƃł���B�����ێs�̃��b�h���X�g�ł͐�Ŋ뜜IA�� (CR)�Ƃ���Ă���B
�����ɂ����鐶���n�̒������ł��邪�A����܂łɂU�����̐����n�����������̂ŏЉ��B�����ێs�U���Y�����≺�D�z���̐����n�ȊO�ł́A������p�̉��Ŕg�̉e�����قƂ�ǂȂ����͐�̉͌������̗Ⴊ�����A���X��⑊�Y��Ȃǂ̑傫�ȉ͐�ł͐����̊m�F���ł��Ȃ������B
�����āA�����n�͒����̖������t�߂Ńn�N�Z���V�I�}�l�L�̐����n�Ɠ������x���i�W���j�ł���A�i�K�~�m�I�j�V�o��n�}�T�W�̐���n����Ⴂ�ꏊ�ł���B�������z�͏\���Z���`�ł͂Ȃ����ƍl�����A�ɂ߂Č��肳�ꂽ�ꏊ�ɂ����������Ȃ����Ƃ��M����B�͌��̏ꍇ�̓t�g�w�i�^����J���A�C�Ȃǂ������Ɍ�����B
�̐F�������n�̍��D�̐F�Ƌɂ߂Ď��Ă���A����Ȃ��ƒT���Ȃ����A���D������Ȃ���̉a����ƂƂ��ɔr�������邽�߂ɁA����ɔr�������c��B�����ڈ��ɒT���Ɨe�ՂɌ����邱�Ƃ��o����B
�����ێs�̎Y�n
�@���D�z��
���D�z���̗V���n�ŁA����ŊC�Ɗu�Ă��Ă��邪�A���傪���S�ɕ܂�Ȃ����߁A�C������������B�C���ɐZ���镔���̓V�o�i���������V���܂�ɐ��炵�Ă���B���̐����n�Ƃ͎�قȂ邪��ɎR������͒W�������ꍞ��ł���B�����ۂ����D�n�ɐ�������B
�A�U���Y����
�U���Y�����̊C�ݗт��A�������ݏo�Ă���ꏊ�ɐ�������B�}�ӂł͊O���w�ʂ͍��F�Ƃ��邪�A�����̌̌Q�̑̐F�͒��F�ł���B����͐����n�̍��̐F�Ɠ����ł��邱�Ƃ���A�ی�F�������Ƃ�����̂�������Ȃ��B
�k���Y�S
�@�]�}��
�@�@��̉͌��ō]�}�p�̓���g�p�̈ꕔ�ŁA�����ۂ����D�n�ɐ�������B
�A�����X��
�����X�Y�̍ł������A�Â��̉͌��̍����ۂ����D�n�ɐ�������B�����ł͉͐���C���ɋ����u���b�N���g���Ă��邪�A���̒��ɗ��܂������D��ɐ���������̂������B
���ˎs
�@�R����
�@�@������͌��̍��ݑ��̍����ۂ����D�n�ɐ�������B
�A������
��{�Y�ɒ������×ǐ�̉͌���ɐ������A�i�K�~�m�I�j�V�o��n�}�T�W�Q�����͂��ɒႢ�����ɐ�������B�����ۂ����D�n�ɐ�������B
���ދn�S
���C��
�����ǐ�̉͌���ō��݂ɐ����B�i�K�~�m�I�j�V�o��n�}�T�W�Q�����͂��ɒႢ�����ɐ�������B�����ۂ����D�n�Ŏ��̉e�ɂȂ镔���Ɍ�����B
�@�^�P�m�R�J���j�i�i�g�Q�J���j�i�ȁj��
�k���Y�����ɂ����镪�z
���茧�����w��@��53�C2001
�����P��
�͂��߂�
�@�^�P�m�R�J���j�i �@Stenomelania rufescens(Ma-rtens,1860)�́A�{�B�i�֓��n���ȓ�j�E�l���E��B�ɕ��z����ŗL��ł���B�����n�͐�̉͌��Ȃǂ̋D����̓D��ŁA�k��20�`80mm�ɂȂ�B�a���̒ʂ�^�P�m�R�̌`�Ɏ��������L�ł���B

�@�{��́A������S�R����b����(1993)�ɂ��ƁA�O�d�Q�A�����V�A���Q�P�A���m�Q�A�F�{�V�A�啪4�A�{��Q�A�������U�A����P�̌v32�ӏ��Ő������m�F����Ă���B�@
�@����AWWF Japan Science Report(1996)�ɂ��ƁA�u��Ő��O�v�Ƃ���Ă���B�����āA�������鐶�L�̎Y�n�͒��������Ȃ��A���m������͌��E�a�̎R���y�c��͌��E���Ɍ��x����͌��E���m�����m�s�|�ԉ������ӁE�F�{���ʖ��s�̗L���C���݁E�������O���s�m�ΊC���ӁE�������p���݂ɎY�n��������x�ŁA���ɂ͏�Ȃ��Ƃ���A30�N�ȏ�O�ɂ͐��L�������Ă������˓��C���h��A���{�C����A���C��ł͋ߔN��ł����Ǝv����ƋL�ڂ���Ă���B
�@�Ȃ��A�L���C�̂������̂����i2000�j�ɂ��ƁA�F�{���ʖ��s�e�r��Ő�ŁA���s����̉͌����L���C�B��̎Y�n�Ƃ���Ă���B
�@�{���ł͖x����s(�Ғ�)�̒��茧�Y�L�ޖژ^�@(1964)�ɑ吣�˒��ł̋L�^�����邪���̌�͂̕Ȃ�����͕s���ł���B
�^�P�m�R�J���j�i�̔���
�@�����ێs�I�����̓s�s�v�擹�H�u���Y�|�I�����v�\��n�ŃE���M�N�̕ۑS��Ƃ��s���Ă���ۂɕς�����L���������B������w�����ق̍��X�ؖҒq����ɓ�����˗������Ƃ���^�P�m�R�J���j�i�ł��邱�Ƃ����������B
�I�����̐����n�͕�40�`100�p����50�����̍a��̊��n�B�ȑO�͊����ł����������ߗ��Ă��A�����̒[���a��Ɏc����Ă���B���̍a��̏ꏊ�ŁA�D�̒��ɔ��ΐ����Ă���{��������B
���k�U���Ǔs�s�v��ۂ��u�ӂ邳�Ǝ��R�̉�v�̗v�]���I�����̊L�ނ̐������������R��w�̕��c�G�������Ɉ˗������B�����̌��ʃ^�P�m�R�J���j�i�͂킸���Ȕ͈͂ɖ�400�̂��������鎖���킩�����B
�k���Y�����ł̕��z
�@�I�����̃^�P�m�R�J���j�i�̐����n�̐����������s���Ă���������Ђ����X��ł��{����m�F���Ă������Ƃ�������������B�������n�Ŋm�F�������̐���30�̑O��ł������B
�@���̉͐�ɂ���������\��������Ǝv���A�k���Y�����̉͐�������B
�@�ȉ��ɎY�n���L���B�@
�͐얼/���ݒn/3�����b�V���R�[�h/�͐쉄���i�Q���͐암�j/�ڎ��ɂ���܂��Ȍ̐��̏��ɋL�ڂ���B
�@���Y��/�����ێs�؋{��/4929-65-23/20km/1000�́j
�A�{����/�����ێs������/4929-46-95/5.2km/50��
�B�����X��/�k���Y�S�����X��/4929-64-79/4.1km/20��
�C�]�}��/�k���Y�S�]�}��/4929-75-60/9.7km/50��
�D������/�k���Y�S������/4929-74-59/4.9km/300��
�E�u����/���Y�s/5029-05-06/11.5km/4��
�ȏ�̂U�͐�Ŋm�F�����B
�@�����n�͂�������ɂ₩�Ȍ��z�ƂȂ����������̏㕔�t�߂ł���B��������̗���ڂɎȂ����ǂ̂悤�ȏꏊ�B�������͊ɂ₩�ȋȂ���̓����̃��V�������Ă���悤�ȏꏊ�ł���B�����ꏊ�ɂ̓C�V�}�L�K�C��J�m�R�K�C�Ȃǂ��������Ă���B
�{��͗c�������x���[�W���[�c���Ƃ��ĉ߂������߂ɊC���̍s�������X���[�Y�łȂ���Ȃ�Ȃ��B��L�̎Y�n�͂�����������傪�Ȃ��͐�ł���B�����X��̎Y�n�͉��̂�����O�̂킸���ȕ����ɐ������Ă����B
�@���������r�I�傫�ȉ͐�̋D����ɐ������D�̒��ɐ����Ă��鎖�������ڎ������ł͊m�F���ɂ����B�܂��A������͂킸�������[�g������\�����[�g���͈̔͂ł���B���̂悤�Ȃ��Ƃ��{��̊m�F��̏��Ȃ��̌����ł��낤�B
�@�^�P�m�R�J���j�i�̐�����͉͐�̉������炷��Ɠ_�ł����Ȃ��A�������̏㕔�t�߂ɂ��ǂ̗l�ȏꏊ���K�v�ł���A���ɔ����ȃo�����X�̏�ɐ������Ă���B
���̂悤�Ȃ��Ƃ��炿����Ƃ������͂ɂ���ł���댯�����ɑ傫���͐�̌@��E�͐ϕ��̏����E���C�ɂ������Ă͍ŐV�̒��ӂ��K�v������B
�@����A�{��̓x���W���[�c�������o�邱�Ƃ��琶������V���ɐ����邱�Ƃɂ�萶���n�̊g�����\���ɍl������B
���@��
�������T��(2000)�D���L���T�i���c�G�j�D�L���C�̊C�̂������̂����D�C���ЁF120-121pp
WWF Japan Science Report Vol.3 (1996)�D���{�ɂ����銱���C�݂Ƃ����ɐ�������ꐶ�����̌���F23pp
�x����s(1964)�D���茧�Y�L�ޖژ^�D���茧�����w��F85pp �D
��S�R����b����(1993)�D�����F30-31pp
���茧�����w��@��48�C1997
�����P��
�͂��߂�
�͌��̟��ւ⊱���̖����A��ݍH�����ɂƂ��Ȃ��ɂ₩�ȌX�������������Ȃ��Ȃ�A�n�N�Z���V�I�}�l�L Uca lactea lactea De haan �̐����n���e�n�ŏ��ł��A�̐��̌������łɎ����Ă���B���̂��ߊ����ł́u���{�̐�ł̂�����̂���쐶�����v�ɂ����Ċ�ɑI�肵�Ă���B
�@���҂͖k���Y�����ɂ�����{��̕��z�n�����Ă��邪�A�����n�������Ă��邽�߁A�����n���l�H�I�ɑ����ł��Ȃ����̂���X�l���Ă����B������R�ł͂��邪�A���֓D���̂Ă��ꏊ�������ƂȂ�A�{��̐������m�F���ꂽ�̂ł��̏ꏊ�Ə��Љ��B ��ł̂�����̂���쐶�����͕ی삪�}���ł���A���̎Q�l�Ƃ��ė~�����B
�@�Y�F�b��11.8�o�E��17.5�o�A���F�b��11.5�o�E�b��17.8�o�B�b�͉��ɒ����l�p�`�B��r�͎��ł͍��E�Ƃ��ɏ��^�ő��́A�Y�ł͒�������������B����̍��E��͂P�F�P�B����͂��G�����A�\�ʂ��������Ƃ��a���̗R���ł���B���p��͌��ŏ����̖������ɒ��ɂ��邭�炢�̓D�̍����������n�ɐE������Đ�������B�������ɂ͍E����o�ďW�c�Ŋ������A�����U�蓮�������A���̂Ƃ��������炪�ǂ��ڗ��B�@�@�@
�@�ώ@�����Ă���Ƒ����U�肩�����đ�����Њd���A�݂��ɑ�������݂��킹�Ă�������̂��悭������B�Ă̔ɐB���ɗY�͔M�S�ɑ����U�莓��U���B�܂��A�����U��Ȃ���̂��㉺�ɏ�������������������邱�Ƃ�����B���������������Ƃ���ɔM�S�ɑ����U��Ȃ��炠�Ƃ���������Ď����̑��E�֗U���B�����Ď����Y�̑��E�֓���ƗY�����E�̊W������B���̑��E�̒��Ō���������Ȃ���B���̍��͗Y�̑��E�̓�����ɂ͊W�����邽�߂̓D���p�ӂ���Ă���B
�@���X�͌���㗑��������܂ܖ�P�����łӉ������A�Ӊ���Ƀ]�G�A�c���Ƃ��ĊC�ɕ��B�]�G�A�c���͔�r�I�L���C��Ńv�����N�g�������𑗂萔��̒E����o�Đ������A���K���p�c���ƂȂ�B�����ă��K���p�c���͂��ꂼ��̐������ւނ��j�����B�{��̏ꍇ�͖�P�����Ŋ����Ȃǂɓ��B���A�����Ő�����������悤�ɂȂ�B
�Q�C�����ێs���̐����n
�@�����ێs�ɂ̓g�m�R���i�Q�����j�E���F���i�����j�E�����i�����j�E���ˁi�R�����j�j�E���F��͌��i�V�����j�E���p�i�Q�����j�E���Y�̂V�����A�k���Y�S�ɂ͍��X��͌��i���݁E�E�݁j�E�����X�������X�Y�E�]�}���]�}�p�����̂R�����̐����n������B
�@�����ێs�ōł����������B���Ă��鑁�˂͖{��̐����n�Ƃ��Ċ��҂������A�D�����������������������Ă��炸�Q�`10�u�قǂ̃R���j�[���R�����Ō�����ɉ߂��Ȃ������B
�R�C�����n�̏���
�@����܂ł̒����Ő����n�͉͌����蓇�̓��]�Ȃǂ̔g�Â��ȏꏊ�����������B����͉͌��Ȃǂ̐����n���Ȃ��Ȃ������ʂł��낤�B
�@�{��̐����n�̊����l�@����ƁA�����n�̏����Ƃ��Ď��̂悤�ȋ��ʓ_���������B
�@�����̎��̖������t�߂ɐ������邱�Ƃ���A�������t�߂��Ȃ��炩�ȊC�݂ł��邱�ƁB
�A�D�̍����������n�Ő[����40�p�ȏ゠�邱�ƁB
�B�l���̂��Ă����܂Ȃ����x�̍d���ł��邱�ƁB
�C�g���Â��ȉ��₩�ȊC��ł��邱�ƁB
�D�����͂��قNjC�ɂ��Ȃ��Ǝv����B
�i���F��͌��ł� BOD�F����8.3 ppm �ő�24ppm �ŏ�3.7ppm�j�i���f�C�I���F�ő�19,400ppm �ŏ�9,900ppm�j�������A�����������̈���ł���Ƒ͐ς���L�@���̗ʂ������Ȃ荻�D�����D�������Ă���ƍl������B�����Ȃ�ƃn�N�Z���V�I�}�l�L����`�S�K�j��}�g�I�T�K�j�̐����n�ւƕω�����ł��낤�B�@�@
�S�C�����n�̑����ɂ��ی�
���p�̉����ɂ��鍌�N���[���Z���^�[�n��̊C�݂ɂ͖ʐϖ�40�u�A�I�X65�̂̃R���j�[�����邪�A�����ێs�`�p���ɂ��p�S�̂ƍ��N���[���Z���^�[�̗��đւ��ɂ�閄���̌v�悪����A�����ꂱ�̐����n�͏��ł���B
�@�������瓌�ɖ�300���̊C�݂ɁA�����R�`�S�N�ɂ����Ďs�̍`�p�������Y���s��V�݂ɔ����q�H�����̟��֓y�����3200�u�i80�~40m�j�������Ă��邪�A���̂����̖�240�u�������̑������D�����ƂȂ��Ă���B���̖�240�u�̍��D�n�ɁA�����P�N�Ԃ͖ڂɌ����鐶���͌����Ȃ������B�������A�����T�N10���ɂ͐��̂̃R���c�L�K�j���ώ@�����悤�ɂȂ����B�����āA�����U�N�T���ɂ̓n�N�Z���V�I�}�l�L�̃I�X�����̊ώ@����A�����V�N�V���ɂ̓I�X75�̂��ώ@�����悤�ɂȂ����B
�{��̐����n�͏����̖������t�߂́A���������D�n�Ƃ������R�E�̔����ȃo�����X�̏�ɐ��藧���Ă���A�{��̕ی�̂��߂ɐl�H�I�Ɋ����̑������s���Ă��蒅�����邱�Ƃ͋ɂ߂ē���ƍl���Ă����B�������A������R�ł͂��������A���֓D���̂Ă��ꏊ�������ƂȂ�A�{�킪�������n�߂��B
�C���Ȃǂ̉e�����A���̊��������ł��Ȃ�����A�V���Ȑ����ꏊ�ƂȂ邱�Ƃ��l������B���̂��Ƃ͐l�H�I�Ɋ���������A�{��̕ی삪�\�ł��邱�Ƃ��������Ă���B���������������C�����̉e���ɂ��i���I�Ɉ��肵�������n�Ƃ��Ċm�ۂł���Ƃ͌���Ȃ����A�ی�̂��߂Ɏ��鉿�l�͏\���ɂ���B�{��̗c�����Ԃ͂P�������ł��邩��A�c�����Y���ł���͈͂Ɋ���������悢�B
�@����̗Ⴉ��A�n�N�Z���V�I�}�l�L�������ł���悤�ɂȂ�܂łɂ́A������R�N�قǎ��Ԃ��������Ă��邽�߁A�����̐����n�����ł���ȑO����v���i�߁A�ώ@�𑱂���K�v������B���N�̒����ł͓��ꏊ���C�����ɂ��Z�H�̉e�����A�����V�N��荻�D�n�̖ʐς��������Ă���悤�ł���B������������ώ@����K�v������B���̂悤�Ȃ��Ƃ���A���S�̂��߈�J���ł͂Ȃ����J���̊��������邱�Ƃ��K�v�ł��낤�B
���Y�̌̐��͑o�ዾ�ɂ��J�E���g�������l�ł���B
���@��
���c���ρi1978�j�J�j�̐��ԂƊώ@�D45�|46�D�@�j���[�T�C�G���X�ЁD����
�O���ˁi1991�j���F���{��^�b�k�ސ}�Ӂi�U�j�@�D�ۈ�ЁD162PP �D����
�����R�N�x�ō����ێs�̊��i1992�j�����ێs�@�������ۑS�ەҏW�D91�|178
Balanophora tobiracola Makino
�i�c�`�g�����`�ȁj�������ێs�����ɂĔ���
���茧�����w��@��48�C1997
�����P��
�@�{�Ȃɂ͖�15��100��قǂ�����A�M�ђn���ɑ����A���̒n���s���������Ɋ���B�a���L�C���c�`�g�����`�̃L�C���͎�����������̒n���Ɉ���ł���B�����̕��z�͋�B�암�E�����ł���A�{��암���܂߂��A��B���C�݂ɕ��z����u��B����蕪�z�^�̐A���v�̂ЂƂł���B
�@���茧�A�����i�O�R1981�j�ɂ́u�����̕��z�n�͒���s�{�͓��������{�сE����s���䗅�R�E�����R�E�F�R�E�O�̉Y�E�ؔ��E����E���c�E�����ˁE�O�d�A���E�e���E���ޑ哇�E�����E�����i��˒��j�E���ʓ��i�ᏼ���j�A�L�쒬�������E�V���ڏ����ł���B�v�A�u�h��̓g�x�����ł����ʁA���ŃV�������o�C�A��Ƀl�Y�~���`�ł��邱�Ƃ����������Ƃ߂��B�ߔN�A�������������Ńn�}�q�T�J�L�Ɋ������̂����������B�v�Ƃ���B
�@���҂͐��ޑ哇���̕S���x�t�߂�1991�N�ɃV�������o�C�ƃg�x���Ɋ��Ă���{������߂Č������A���̐��̑����ɋ������B�����ł͔�r�I�Â����x�̍����ꏊ�ɑ������炵�Ă����B�哇����k�̊C��ɍ����ێs������������B�����ɂ̓��N�^�`�o�i�E�T�c�}�T���L���C�E�n�}�T���g���C�o���ƂR��̖k���̐A�������炵�Ă���B���̓��ɂ��{�킪���z���Ă���\���������Ǝv���A1994�N���璲�������Ă������A1996�N12���R���A���łɉԊ��͉߂��Ă������A�Q�����Ńg�x���Ɋ��Ă��鍇�v�W�{��T�����Ƃ��ł����B����ɂ��A����܂ł̐��ޑ哇�ɑ��荕�����k���ƂȂ����B
�@���ޑ哇�ł͓��H���痣�ꂽ�т̒��ł��������A�����ł͓��H�ۂ̗ѓ��Ō���ꂽ�B���������Ȃ芣�������ꏊ�Ŋ��I�ɂ͑O�҂Ƃ͂��Ȃ�قȂ��Ă������A�̂����������̂ł������B����n�̓^�C�~���^�`�o�i�D��̒�ؗтŁA�N���L�E���u�c�o�L�E�q�T�J�L�E�V�������o�C�E���`�m�L�E�J�N���~�m�Ȃǂ������A�я��ɂ̓z�\�o�J�i�����r�����炵�Ă����B
�]�k�ł��邪�A��������q��V���{�A���}�ӂɂ́u�c�`�g�����`�͒n���s����g�����`�����v�Ƃ���̂ŁA�哇�ō̏W�������̂Ŏ��݂����A�{�킩��̓g�����`�͂ł��Ȃ������B�܂��A�}�łł͖{��ƃ����E�L���E�c�`�g�����`������ւ���Ă��邱�ƂɋC�t�����B
���@��
�k���l�Y�E���c���i1976�j�D���{���F�A���}�ӁD316-317. �ۈ�ЁD����
�����O���i1996�j�D��B����蕪�z�A���F��`�C�\���C�N���G�A�����ށC�n���D47:119PP
�O�R�O�Y�i1981�j.���茧�A�����D���茧�����w��D47PP
�q��x���Y�i1995�j�D��������q��V���{�A���}��.61-62�D�k����.����
�V�I�A�����{ Asclepios shiranui Esaki,1924�i�A�����{�ȁj�ƃV���E�~�A�����{ Halobates matsumurai Esaki,1924�i�A�����{�ȁj�茧�����ێs��\�㓇�Ŋm�F�����B
���茧�����w��@��53�C2001
�����P��

���V�I�A�����{��
�����̃A�����{�ނ͋��ɉ��ݐ��ł���A�����n�悪�����Ă���A��ł̋��ꂪ���������̃��b�h���X�g�ł̓V���E�~�A�����{���Ŋ뜜�T��(���茧�ł����l)�A�V���E�~�A�����{���Ŋ뜜�U��(���茧�ł͇T��)�ɑI�肵�Ă���B
�@��\�㓇���̔g�̉��₩�ȓ���]�ňȑO����A�����{������̂͒m���Ă������A��N�X����10���ɂR��̃A�����{�����߂č̏W�����菗�q�Z����w�r��P�������ɓ�����˗������B���̌��ʃV���E�~�A�����{�E�V�I�A�����{�E�P�V�E�~�A�����{�̂R��ł��邱�Ƃ����������B
�V���E�~�A�����{�E�V�I�A�����{�E�P�V�E�~�A�����{�̂R�킪�A�܂��V�I�A�����{�ƃV���E�~�A�����{�̂Q�킪�����Ɍ�����ꏊ�A�V���E�~�A�����{������������ꏊ�Ȃǂ܂��܂��ł��邪�A��������D��Ő��[���g�����₩�ȓ���]�Ɍ�����B
�@�V�I�A�����{������]�̉����ł̂݊ώ@���ꂽ�̂ɑ��V���E�~�A�����{�͐��[��������x�������]�̓�����◣�ꂽ�ꏊ�ł�����ꂽ�B�܂��A�V���E�~�A�����{�ɔ�ׂăV�I�A�����{�̐����ꏊ�͂��Ȃ菭�Ȃ������B
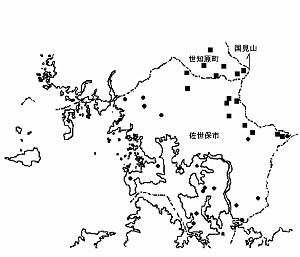
�@����܂Ŋm�F�����ꏊ�́i�\�P�j�̒ʂ�ł���A��\�㓇��т̓V�I�A�����{�ƃV���E�~�A�����{�̋M�d�ȎY�n�ƍl������B��N�̒����͋�\�㓇�̈ꕔ�ł����������ێs���Ɍ����Ď��{�����B�k���C��≈�݂ɂ��������Ă���ƍl������̂ŁA������L���n��ł̕��z�������s���Y�n�m�ɂ���K�v�����낤�B
�@�@�@�i�\�P�j
|
�a�@�@�� |
�Y�@�@�n |
�R �����b�V���R�[�h |
|
�V�I�A�����{ |
�g�R�C�� |
4929-74-28 |
|
�V�I�A�����{ |
���Y�� |
4929-55-64 |
|
�V���E�~�A�����{ |
���� |
4929-74-26 |
|
�V���E�~�A�����{ |
��~�ؓ� |
4929-74-38 |
|
�V���E�~�A�����{ |
�g�R�C�� |
4929-74-28 |
|
�V���E�~�A�����{ |
�g�R�C�� |
4929-74-38 |
|
�V���E�~�A�����{ |
�㏬���� |
4929-74-29 |
|
�V���E�~�A�����{ |
�g�m�R�� |
4929-74-29 |
|
�V���E�~�A�����{ |
���̓� |
4929-55-94 |
|
�V���E�~�A�����{ |
���Y�� |
4929-55-64 |
�Ӂ@��
�@�{���܂Ƃ߂�ɂ�����A���n�̒����y�щ��ݐ��A�����{�̓�������Ă����������A�r��P������Ɍ�������\���グ�܂��B
���茧�����w��@��55�C2002
�����P��
�@�@
���茧�����ێs�Ɩk���Y�S���m�����̃��}�A�J�K�G���ƃj�z���A�J�K�G���̕��z�ɂ���
���茧�����w��@��53�C2001
�����P���@�@�@�@�@�@
�͂��߂�
�@���҂�1989�N���猧�k�n��ɂ����闼���ނ̕��z�������s���Ă��邪�A�J�K�G���Ȃ̃j�z���A�J�K�G�� Rana japonica Gunther�ƃ��}�A�J�K�G�� Rana ornativentris Werner�̍����ێs�Ɩk���Y�S���m�����ɂ����镪�z���قڂ��ނ��Ƃ��ł����̂ŕ���ƂƂ��ɁA�ɐB���Ɋώ@�ł����s����������B
�������@
��ɔɐB���Ɨc����������12�`3���ɂ����āA�ɐB�n�̐��c����r�ŗ���E�c���E���̂�T�����B����ł̊m�F�͍L�����j�E�x������(1974)�ɂ����@�ŁA���̂悤�ɍs�����B�V��������̏ꍇ�j�z���A�J�K�G���̗���́i�}1�j�̂悤�Ɏ�Ŏ����グ�邱�Ƃ��o���邪�A���}�A�J�K�G���́i�}�Q�j�̂悤�Ɏ����グ�邱�Ƃ��ł��Ȃ��B

�i�}1�j�j�z���A�J�K�G���̗���@�@�@�@�@�i�}2�j���}�A�J�K�G���̗���@�@�@�@�@�@�@
��Ԃ̒������s���Ɨ���̉��ɃI�X������ꍇ�������A�m���Ɏ킪����ł���B�Ȃ��A����̏ꍇ�ɂ͗c���̍��ɍēx�ώ@�ɏo�����Ă��m���ɓ��肵���B�c���ł̓j�z���A�J�K�G���̔w�ʂɈ�̍����i�}�S�j������̂ɑ����}�A�J�K�G���i�}�R�j�ɂ͖������ƂŔ��f�����B
 �@
�@
�i�}4�j�j�z���A�J�K�G���̗c���@�@�@�@�@�@�i�}�R�j���}�A�J�K�G���̗c��
�����}�A�J�K�G���ɂ���
���}�A�J�K�G���͎R���i1967�j�ɂ���Ėk���Y�S���m���������R�ł̐�����������Ă��邪�A���̌�͖̕��������B���҂�1990�N�ɐ��m�����Ŗ{����m�F���Ă���B
���}�A�J�K�G���͍����ێs�����̕W��100m���瓌�k���̕W��450���ɂ����ĂƐ��m�����̕W��240�`510m�t�߂ɂ����ĔɐB�n������A�Y���n�͐��c�₻�̐��H�Ɛ��r�ł���B
�@�ɐB�s���́u�h���N���a�v�ƌĂ�鏬�J�̍~��悤�Ȓg������ɍs���邪�A���҂̊ώ@�ɂ��Ƒ����͎��x��������Ԃ̋C�����T���ȏ�̎��ɍs����B�Q�����{�`�R���̓j�z���q�L�K�G���̔ɐB���Əd�Ȃ�B�Ƃ��ɂ͈ꏏ�ɏo�Ă����q�L�K�G���ɂ����ڂ���̂�����ȂNJώ@���Ă��ĖO���Ȃ����̂ł���B
���m�����̕��͌��r�ł́A�Q���̒g��������������������ɗ��r���ɐ��\�C�P�ʂɏW�܂芈���ɉj���Ȃ���I�X���m�����ڂ�����A���ꂽ��̍s�������Ȃ��琷��ɖ��̂�1992�`1993�̂Q�N�Ԋώ@���ꂽ�B1992�N�ɂ͂R�J���ō��킹��235�C���̃I�X���W�܂肱�̂悤�ȍs�����ώ@���ꂽ�B
�@���̍s���͖�Ԃł͂Ȃ����Ԃł���A�T�`10���������炭�x�ށB���̂悤�ȍs�����Â��Ȃ�܂ŌJ��Ԃ��ꂽ�B���������̊ԃ��X�̎p�͐��C���������Ȃ������B���̎��̓��₩�Ȗ������͍����ێ��R�K�C�h�u�b�N�i1996�j�ɓY�t�̂b�c�ɔ[�߂Ă���B���̔N�͂��̌�Ɋ��g���P���������܂�ɐ���ł������}�A�J�K�G�������\�C���B���̌�̃h���N���a�ɂ͐�Ɏ��̂ɑ����̃I�X�����ڂ��Ă���̂�����ꂽ�B�����Ȃ����̂����X�Ǝv�����̂��낤�B���т������R�̈�ʂ�������ꂽ���̂ł���B
�@�Ƃ��낪��������Ă����u���b�N�o�X�̐����ƂƂ��ɂ��̒r�ł͂��̂悤�ȍs���̊ώ@�͂ł��Ȃ��Ȃ�A�{��̗���i�ő��138����j�A�̐��Ƃ��Ɍ��������݂͏\���̗��������Ȃ��Ȃ����B�������ꏊ�Ō�����q�L�K�G���̗��ɂ͑傫�ȕω��͂Ȃ��B
�@�{�킪�u���b�N�o�X�ɕ�H����Ă��錻���ڌ��͂��Ă��Ȃ����A�����̌����̓u���b�N�o�X�ɂ���H�Ǝv����B�Ȃ��A�����ێs�،����i�W��100m�j�ɂ͗B�ꗼ�킪�ɐB���鐅�c������B
���j�z���A�J�K�G���ɂ���
�j�z���A�J�K�G���̕��z�̓��}�A�J�K�G����肩�Ȃ�L�����A�����ێs�������琼���ɂ͕��z�n�����Ȃ��A���m�����ɂ͐������Ă��Ȃ��B
�{��̎Y���͒�n�̐��c�₻�̐��H����ł���A���}�A�J�K�G���̂悤�ɗ��r�ł͊ώ@�������Ƃ��Ȃ��B
�@��������̂悢�g�����ꏊ�ł�12������Y���������A�R���܂ő����B�ɐB�n�͊C�݂���W����250m�܂łł������B
�ɐB�s���̓��}�A�J�K�G������Ԃ̋C�����������ɑ���10�����z����ꍇ�������B�P�C�̃��X�ɂQ�`�R�C�̃I�X�����ڂ���Ȃǂ̍s���͌��ĂĊy�����B�����̓��}�A�J�K�G���̗l�ɑ傫���Ȃ��Â��ŁA�������𗧂ĂȂ���Ε������Ȃ��ꍇ������B
���@�z
�@���ꂼ��̕��z�n���R�����b�V���R�[�h�Ɓi�}�W�j�Ɏ����B����������͐��̂̊m�F�ł���ȊO�͔ɐB�n�B
�����}�A�J�K�G��
�@ �����ێs�F�˃��q��4929-66-33,34�A����R���z4929-66-13���A������4929-66-14,24�A���B���x4929-66-04���A�،���4929-56-89,��97���A�M�،���4929-66-50,61�A�S�쒬4929-66-15
�A���m�����F�J���4929-66-82�A�㌴��4929-66-81�A���͌��r49-29-66-72�A�Ԗ؏��4929-76-02
�����R4929-66-74,��84��
���j�z���A�J�K�G��
�@ �����ێs�F�I����4929-65-43�A�|�Ӓ�4929-65-34�A���c��4929-65-16�A���蒬4929-56-96�A4929-66-07�A�،���4929-56-89�A���̉Y��4929-55-45�A�U���Y��4929-55-44�A�j������R�E�c���E�{�Y4929-56-21,31�A�앗�蒬4929-56-04�A�{�Ò����Y4929-46-84�A������4929-56-27�A�j������4929-46-80,90�A���D�z��4929-55-66���A��Ӓ�4929-55-58
�i �}�W�j�����}�A�J�K�G�� ���j�z���A�J�K�G��
�I����
�@�ߔN���c����������Y���n������������ޏꐮ���ɂ�肻��܂ŎY���n�ł��������H���R���N���[�g�����ꐅ�����܂�Ȃ��Ȃ����萅�c����������悤�ɂȂ����B�܂����m�����̕��͌��r�̂悤�Ƀu���b�N�o�X�̕����ɂ��Y���������������ꏊ������Ȃǔނ�̔ɐB�ɑ��鈳���������Ă���B
�� ��
1997�N����͍����۔Ń��b�h�f�[�^�u�b�N�ҏW�ψ��̗V�����q����A�哇�ʍO����A���茪��Y����A�Óc���q�v����ɂ͊������ꏏ�ɒ��������Ă��������܂����B�{���s���ɂ����肨��\���グ�܂��B�@
���@��
�R���S�j�i1975�j.���茧�̐����D���茧���ȋ��玖����
�L�����j�E�x������(1974)�D�j�z���A�J�K�G���ƃ��}�A�J�K�G���̗���̔�r�D��`�D28.3:108-111
�����P��(1990)�D���茧�ɂ����郄�}�A�J�K�G���̍Ĕ����D���茧�����w��D���茧�����w��F41-43
�ܓ�����p���������j�z���~�c�o�`
�@�v�u�y�m�j
�@�������{�ŗL��̃j�z���~�c�o�`�������n�߂ĂQ�O�N�]��ɂȂ�B���̂Q�O�N�̊Ԃɏ��������R�E����j�z���~�c�o�`�̐������������Ă��Ă���悤�Ɏv���ĂȂ�Ȃ��B������̎����Ɏ��̎d�|�����u�҂����v�ɓ���V�Q�̐������Ȃ��Ȃ��Ă��Ă���̂ł���B�����͒n���̉��g���ł��낤�Ɣ��R�ƍl���Ă����B
�@��N�̏t�A���Z�̓�����̂���A��]�҂����Ōܓ����s���s�Ȃ����B���]���̓����̋߂��ɍ̉Ԃ��炫����Ă����B���̓j�z���~�c�o�`�̎p��T�������S��������Ȃ������B�߂��ɗ{�I�Ƃ�����炵���Z�C���E�~�c�o�`�̎p�͂������B���]�`�̋߂��Ƀ����Q�c���������̂Ńo�X���~�߂Ă��炢���ׂ����A��͂茩����Ȃ������B�Q�O�N�قǑO�ɂ͌ܓ��ɂ̓j�z���~�c�o�`�������̂ł���B�V���L���Ō����������������B�t�ɁA�R�ɑ������d�|���Ă����A�H�Ƀ~�c�o�`��S�ďĂ��E���Ė����̂�b�ł������B�u����Ȃ��Ƃ�������n�`�����z�v�Ǝv�����L�����������B
�@���N���{�Ăѕ��]���ɍs�����B����͕��]���̐^�ɂ���R���f�Ï��̋{�菺�s��t����A�ܓ��Ƀj�z���~�c�o�`�����������̂ŋ��͂��Ă���Ƃ̗v��������A�Ē����ɏo�������̂ł������B�̉Ԕ��ɂ͂܂������j�z���~�c�o�`�̎p�͂Ȃ������B��ł̌����͔���Ȃ��܂܁A���͂���ĕʂꂽ�B
�O����{�A�~�̍炭�Ƃ��A���͏�ܓ��������Ȃ̂��ǂ��������ɏo�������B�S�����o�C�N�ő����������A���Ƀj�z���~�c�o�`�͂P�C�������ł��Ȃ������B�~�ɂ͏����ȃA�u�����Ă������̉Ԃɂ͂ǂ�ȍ��������Ă��Ȃ������B�s�v�c�������̂͐l�Ƃ̖T�̔������Ɏ��𒅂��Ă��鎖�ł������B���͏o��V�l���Ƀj�z���~�c�o�`�̏�����q�˂��B�قƂ�ǂ̐l����u�̂͂������ŋ߂͌������Ȃ��v�Ƃ����������Ԃ��Ă����B�����āu���̒킪�̎����Ă����v�Ƃ����l�ɏo��A�L�쒬�������̉F��v�v����ɍs���������̂ł���B
�@�u�\���N�O�܂ʼnƂ̗��Ŏ����Ă����B�ޗǔ��̐l����ꔠ������̂��n�܂�ł������B�i�X������8���ɂȂ������A����Ȍ㌸��n�߂R���ɂȂ����B�Ƃ��낪���̔N�A�n�`�����͌��C���Ȃ��A�������߂Ȃ������B�X�Y���o�`�ɏP��ꂽ���n�`���s���Ő킦���S�ł����B����Ȍ㎩�R�E������p���������v
�@���ꂪ�F��v�v����̐����ł���B�����͔���Ȃ����A�������̔N�͔~�C���A���Ȃǂ̉Ԃ������������悤���Ƃ̂��Ƃł������B�ܓ��ʼn����N���邢�͉��\���N�����������Ă����j�z���~�c�o�`���\���N�O�ɓˑR��ł����̂ł���B���ɂ������Ă���l���������ǂ����q�˂��Ƃ���A�m���Ă������ł͂Q�l�������S���Ȃ�ꂽ�Ƃ̂��Ƃł������B�������c���Ă����猩���Ă��炢�����ƌ��������A���͂⋀���ʂĂĎc���Ă͂��Ȃ������B����������u���Ă����ꏊ�ɂ͈ē����Ă�������B
�@�\���N�O�Ƃ����͎̂��ɂ��L��������B���̎����Ă���2�O�Q�قǂ��S�Ď��ł����N�Əd�Ȃ�悤�ł���B���̔N�͒g�~�ŁA�n�`�͉Ԃ��Ȃ��̂ɔ�щ��A������������B�����ďt�ɂȂ��Ă��~����̉Ԃ��قƂ�Ǎ炩�Ȃ������B���I���w�ǂȂ��A�����̌Q���~�J������Ȃ������B�c�������������X�ɓ|��A�����ɃX�Y���o�`������A�c�����S�Ă̌Q�Ɏ~�߂��h�����B
�@���͂��̔N�A�j�z���~�c�o�`�͓��{�����ł����Ǝv�����B�Ƃ��낪���N�̕��I���ɁA�ǂ����Ő����c���Ă����Q�̕��I�Q���A���ł�����̋��ɂQ�Q�������̂ł���B���ݎ��͂S�O�Q�����Ă��邪�A����͂��̂Ƃ��̂Q�Q����n�܂������̂ł���B
�@�ܓ��ɂ������ł̌�����T��ׂ��A���͌܌����{�Ăя�ܓ��ɓn�����B�ł̉Ԃ��炭�����ł���B�㗤���Ē����������������C�������B��d�ɂ��A�Ȃ�R������ƁA�Έ�F�ł���B��B�{�y�ł͒ł̉ԂŎR�S�̂������Ȃ��Ă��鎞���Ȃ̂ł���B����w�̑��т��R�S�́A����܂Ői��ł���B���̐���w�̊Ԃ̏��X�ɒł̔����Ԃ�����B�O��͐F�������Ȃ̂ŋC�Â��Ȃ������B�G���Ȃ��Ȃ�A���ꂪ���ݏo���H���̖����͊����Đ�ł����̂ł���B
���͍Ăуo�C�N�őS�������ĉ�邱�Ƃɂ����B���̓x�͈ꔑ���Ă̒����ł���B����ɂ͂Q��ނ̃}���n�i�o�`�����Ă����B�Ƃ̎���ɐA���Ă�����x�̉ʎ��ł���}���n�i�o�`�ŊԂɍ����B���̔~���������Ă����B���͉ߑa�n�ő�K�͔_�Ƃ������Ȃ��l�͂��Ȃ��̂ŁA�~�c�o�`�s�݂̖�肪�\�ʉ����Ă��Ȃ��̂��낤���A�����A�Z�ނ��K�͂ɍ������A��ʎ������o�c���悤�Ƃ�����A����͕s�\�ł��낤�B�����Z�C���E�~�c�o�`�����悤�Ƃ��Ă��A�����~�c�o�`��{���Ȃ��̂ŁA�{�I�Ƃ͗��Ȃ��B�i���ݕ��]���ɂ͖k�C���̗{�I�Ƃ������Ă��āA���Ăɑ�^�g���b�N�R��Ŗk�C���ɉ^�Ԃ����ł��邪�A�~�J�Ɠ~�ɂ͍������ŋ��a�����Ȃ���Ȃ�Ȃ��B���ܓ��ɂ͗{�I�Ƃ͓����Ă��Ȃ��B�j
�@���̉ߑa�n�ɓ��H�Ԃ����͒��菄�炳��Ă���B����w���o�����߂̗ѓ��ł��낤�B�Ζʂ��ʂ��č�����K�[�h���[���t���ܑ̕����H�ł���B������C�݉����ɖk�サ�Ă݂����A�ޖ��p�Ɏ���܂łP�l�̐l�Ԃɂ��P��̎Ԃɂ��o���Ȃ������B����ȂƂ���Ŏ��̂��N��������C�Â��Ă��炦�Ȃ��Ǝv���Ƌ��낵���Ȃ����B�R���������P�C�^�C���ʂ��Ȃ��B����ȂƂ���������̎Ζʂ͌o�ϗтŕ����Ă���B�G��蕥���Ă̑��т��Ȃ��ꂽ�킯�ł���B�������}�ł����Ԕ�������Ȃ��ŕ��u����Ă���炵���A�т̒��͂ǂ������ʂ��Ȃ��B�L��Ȓn���l��s���ŊǗ����ł���͂��͂Ȃ��B�o�ϗт��o�ϓI���l�ݏo���Ȃ��̂ł͂Ȃ��̂��B
�@����ǂ��납�A�G���Ȃ��Ȃ�A�~�c�o�`�����Ȃ��Ȃ�ΎG�̎�������A������^�ԏ��������Ȃ��Ȃ�B�₪�Čo�ϗт͑䕗�œ|��A�R�͍r�p����B�C�ɗ����J���͉h�{���R�����Ȃ�A�����l��Ȃ��Ȃ�ł��낤�B�H���A���̊̈�����悤�ł���B���Ƃ����đ����G�̐X�����A�j�z���~�c�o�`�������Ȃ��Ɛl���Z�߂Ȃ����ɂȂ鋰�ꂪ����B
�@�{���t�Ƃ̖͉ʂ��������ɂȂ��B�H�ƕs���̓y�n�ł͌��ǂ͎��ł���ł��낤�B
�@��ܓ��ɒ����ɍs���O�́A��ł̌�����T�邾���łȂ��A���̌Q���ڂ��āA�ܓ��Ƀj�z���~�c�o�`�������邱�Ƃ��ł��Ȃ������ׂ������������B�����āA�k���ڔ����̔Ԋx�Ȗk�ɂ͑��т��Ȃ���Ă��Ȃ����Ƃ������B���܂�ɎΖʂ̌X�������A�A�яo���Ȃ������̂ł��낤�B�j�z���~�c�o�`�������Ă����邩�ǂ����A�Ԋx�̖k�Ζʂɂ���吅�n��̎G�̐X�߂Ȃ���l���Ă݂����A��͂�ʐς�����Ȃ��B�Q�C�R�Q�ł͋ߐe���Ŗłт�B2�O�Q�ȏ�ł͉ߖ��ɂȂ�B����ɂ��̎Ζʂ͖k���ɖʂ��Ă���A�~�͗₽�������C���琁���t����ɈႢ�Ȃ��B
�@�ܓ�����A���Ă��āA�����̎�����o�ϗт̊����̊ϓ_����ώ@�������Ă݂��B�����ċ������B�ܓ��قǂł͂Ȃ����A���Ȃ�̕������o�ϗтɕ����Ă���ł͂Ȃ����B
���������Ă���u���{�I������v�̕ł͑�G����������{��̒ŗt��������N�j�z���~�c�o�`�����ł����Ƃ����������Ă���B���{�ł͕ƒn�A�ߑa�n�قǑ��т��i��ł���B
�U���Q�T���A���͓V���ɂ������ɍs���Ă݂��B���т�����قǐi��ł��Ȃ��̂����āA���̓��͑��v���ƒ��������B�����Ƀ��N���W�̉Ԃɓ��₩�ȉH�����������Ă���̂��������B
���N�͏t�̖K�ꂪ��N���Q�T�Ԓx��A�~�͎�ނɂ���ď������J�Ԃ̓������Ⴄ�̂ł��邪�A���N�͈�x�ɍ炢���B�����ăj�z���~�c�o�`�̕��I�J�n���Q�T�Ԓx��A���I�������Ȃ������B���I�������Ȃ����Q�͑S�̂̂S���̂P�ŁA�������w�ǂ��P��ł������B��N���ƂR���̂Q�����I���A�Q�C�R��s���B�j�z���~�c�o�`�̕��I�قǎ��R���̏�Ԃ𐳒��ɔ��f������̂͂Ȃ��B���タ����Ƃ����C��ϓ��Ńj�z���~�c�o�`�͑S���I�K�͂œˑR��ł��鋰�ꂪ����Ǝv���Ă���B���g�����i��ł���̂ŁA�ǂ�ȋC��ϓ����N���邩�\�z�ł��Ȃ��B��Ŋ뜜��Ɏw�肷��K�v������̂ł͂Ȃ����낤���B
 �@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@
�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@
�L�싽�̈��x�̎R���ŏ�ܓ��̓T�^�I�ȑ��т̏@�@�@�@�@�@�@�@�@�@�@�@�R�̏�܂œ��H���ł��Ă���i���V�i�C�ɖʂ����������ӂ�j�@�@
���}�A�J�K�G���̐V�Y�n�i�����X���j
�����@�P��
�j�z���A�J�K�G���͌����ɍL�����z���Ă��܂������}�A�J�K�G���̕��z�͍����ێs�Ɍ����A����܂ŎO�͓����A�������A�������A�M�ؒ��A���m�����̎R�ԕ��A�g�䒬�ܑ��x�R�[�܂ŕ��z���Ă��邱�Ƃ��킩���Ă��܂����B
����̕��z������1���`3���̔ɐB�����ł��K���Ă��܂��B���̊��Ԃ͎Y���ɏo�Ă������̂◑��A�c������������Ԃ�����ł��B�Y���ꏊ�Ƃ��ăj�z���A�J�K�G���͐��c�␅�̗���Ȃ����H�𗘗p���܂����A���}�A�J�K�G���͂����ɉ������r���g���܂��B
��N�A�����X���̒����ŗ���̗���Ɨc����������܂����B����ɑ������N�ɂȂ萬�̂�������A�������m���ɂȂ�܂����B
���}�A�J�K�G���̂���܂ł̐����n������ƈ��̎��R�т̉���R�ԕ��ł�����������x�W��������ꏊ�ł����B�������A�����X���ł͊C�݂���1�L���قǂ̂������A�W��30������2�̏ꏊ�Ō������Ă��܂��B���̂����̈�A���U�i�Â�j�ł͐��c�̐��H�Ƀj�z���A�J�K�G���Ɠ����ɎY�����s���Ă��܂��B
����܂ł��،����ł͗��킪���݂���ꏊ������܂������A�W����120�����ł����B�����X���̎Y���n�̓��}�A�J�K�G���Ƃ��Ă͍ł��W�����Ⴂ�ꏊ�ɂȂ�܂��B
�����X���̒n�`������ƁA���n���w�ǖ����C�߂�����c��ڂ�����A�����Ē����ɎR�n�ւƂȂ���A���R�т̉�����\�L���c����Ă��܂��B�������A�����R���������s�����Ȃ����߂ɐ��c�̈ʒu���Ⴍ���ꂪ�A��n�ŎY���i�ō���150���j����v���ƂȂ��Ă��܂��B
����Ƃ��ɎY���n�̊������Ă݂�ƁA�߂��ɐ����ꏊ�Ƃ��Ă̎��R�т�����ꏊ�ł��B�������A���}�A�J�K�G���̕������L�����R�т̂���ꏊ�Ɍ�����悤�ł��B�����āA�ǍD�ȍ̉a�ƎY�������������ΐ����͕W���ƊW�Ȃ��킾�Ǝv���܂��B
����A�j�z���A�J�K�G���͊C�߂��̐��c�ɂ��Y���ꏊ������܂��B�������߂��ɗǍD�ȗт����邱�Ƃ������ŁA�Y���n�̍ł��W���������ꏊ�͔��⒬�̕W��150�����ŁA�W��100���ȉ����w�ǂŕ��n���D�ނ悤�ł��B
���̂悤�ɁA�j�z���A�J�K�G���̕��z�͎R�n�ɋy�Ԃ��Ƃ͂���܂����}�A�J�A�K�G���͐��������������Ă���A���n�ɂ��������\�ł���A���̂��Ƃ��l����ƃ��}�A�J�A�K�G���͗ђn�J���ɂ��R�ԕ��֒ǂ����ꂽ�̂ł͂Ȃ����Ǝv���܂��B�ђn�����ɍL���c���Ă������͗���Ƃ������ꏊ�Œ��ǂ��Y�������Ă����Ǝv���܂��B
 �@�@
�@�@
�@�@�@�@�@�@�I�X�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@���ځ@�@